Ciò è vero, poiché tutti i
vertebrati, ciclostomi inclusi, possiedono del materiale più o meno rigido
all’interno del corpo, con funzione di sostegno. Tuttavia questa è una
caratteristica dei cordati e non solo dei vertebrati, perché anche l’anfiosso
possiede una sorta di scheletro cartilagineo interno con funzione di sostegno,
la notocorda.
Ciò che distingue invece i
vertebrati dagli altri animali dotati di un sostegno interno è il possesso del
cranio, di tessuto mineralizzato a protezione del cervello e degli organi di
senso della regione cefalica.
 |
| Apparato scheletrico di missine (alto), lamprede (centro) e condritti (basso). In blu il tessuto cartilagineo, in verde la notocorda, in giallo il tipico tessuto fibroso che avvolge il cervello e la notocorda, tipico delle missine |
E’ vero, se escludiamo le
missine, gli altri vertebrati hanno uno scheletro con vertebre ben definite e
una rigidità maggiore rispetto a quella dell’anfiosso, ma siccome le missine si
sono dimostrate vertebrate a tutti gli effetti (vedi qui), ritengo sia giusto
porre maggiormente l’attenzione sul cranio, che poi è la vera caratteristica
che fa la differenza nello sviluppo dei vertebrati.
Lo scheletro (cranio + serie
dorsale + cinti + scheletro appendicolare) è la parte più importante dei
vertebrati per chi studi quelli fossili, visto che nella maggioranza dei casi è
la sola cosa che rimane nel processo di fossilizzazione.
Siccome lo scopo di questa
serie è aiutarci a capire come possiamo studiare l’evoluzione dei vertebrati e
degli gnatostomi attraverso lo studio dei fossili, riconoscere le varie parti
dello scheletro è a dir poco fondamentale.
Oggi però non voglio
parlare delle singole parti che compongono lo scheletro, ma del materiale che
rende questa struttura anatomica così particolare.
Lo scheletro dei vertebrati
può essere in realtà diviso in due parti molto diverse tra di loro, per
composizione, struttura e funzione.
Da una parte abbiamo lo
scheletro interno, o endoscheletro,
che compone il neurocranio (che contiene il cervello) e lo splancnocranio, che
consiste in mandibole, osso iode e archi branchiali (dunque anche le ossicina
dell’orecchio interno negli amnioti).
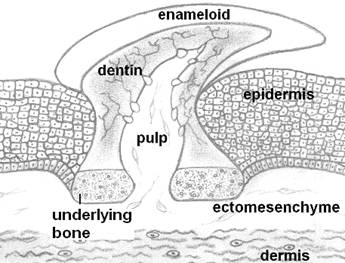
Esternamente vi è invece l’esoscheletro, o scheletro dermico, che compone ad esempio le ossa più esterne del cranio (es. frontale, parietale, opercolare, etc..), i denti e le scaglie.
L’esoscheletro è formato da diversi tipi di tessuti, alcuni tipici di
specifici gruppi altri che invece si trovano nella maggior parte dello
scheletro esterno dei vertebrati.
L’esempio più classico è la
dentina, materiale molto comune formato da cristalli di idrossiapatite. La dentina è molto comune all'interno di gnathostomata: essa infatti ricompre i nostri denti ma è anche, ad esempio, la parte più esterna della
pelle degli squali, che di fatto è formata da piccolissime scagliette a forma
di denti, composti appunto da dentina.
Dello stesso materiale è
composto il tessuto osseo membranoso, che altro non è che una miscela di minerali
come calcio, magnesio e fosfato, unito a una matrice di collagene (a dare appunto
idrossiapatite). Come la dentina, esso si forma dal tessuto dermico ed è
diverso dal tessuto osseo dello scheletro intorno come struttura e origine. Per
la precisione, la dentina si origina dagli odontoblasti mentre il tessuto osseo
membrano dagli osteociti.Quando queste cellule hanno compiuto il loro lavoro, esse possono morire o sopravvivere rimanendo inglobate all'interno dell'osso. In questo ultimo caso si forma tessuto (dentina o osseo) cellulare, altrimenti tessuto acellulare.
Altro tessuto dello
scheletro esterno dei vertebrati è lo smalto, tessuto ipermineralizzato ma non
composto di collagene, che viene prodotto dall’epidermide. Esso è presente solo
negli osteitti, mentre una variante dello smalto, detto “enameloid” si trova ad
esempio in vari gnatostomi, fossili e viventi, ma si differenzia dallo smalto poiché esso non
è prodotto esclusivamente dall’epidermide ma dall’interazione tra questa e il
tessuto sottostante (ectomesenchima). A differenza di ciò che viene insegnato anche in moderni corsi di anatomia, lo strato che ricopre la dentina nei dentelli epidermici degli squali è formato da enameloid, e non da vero smalto.
Per quanto riguarda la
condizione primitiva dell’esoscheletro degli gnatostomi, fino a poco tempo fa
si pensava che essa fosse rappresentata da piccole scaglie di tessuto dermico,
come presente in condritti, acantodi e alcuni agnati, nonostante i placodermi possiedano
un esoscheletro formato da grandi placche dermiche. Tuttavia, queste non venivano
ritenute omologhe alle ossa degli osteitti e la condizione primitiva per gli gnathostomi era considerata micromerica, con un'esoscheletro formato da piccole scaglie. Da qui, gli osteitti avrebbero poi
sviluppato una condizione macromerica, in cui le varie scagliette si sarebbero
riunite a forma le grandi unità dermali (es. opercolare, nasale,
frontale, etc.) che ancora oggi compongono lo scheletro di tutti gli osteitti,
uomo compreso.
Tuttavia, la recente
scoperta di un placoderma con condizione macromerica e divisione delle ossa
della mandibola come quella degli osteitti, ha fatto capire come la presenza di
ossa dermali macromerica in placodermi e osteitti sia omologa, e che quindi la
condizione micromerica dei condritti e degli acantodi è derivata e non
rispecchia quella presente nell’antenato comune dei crown gnathostomi. Ne ho parlato
più approfonditamente (qui)
L’endoscheletro comprende neurocranio, splacnocranio, vertebre, cinti
e scheletro appedicolare. Ognuna di queste parti ha una sua storia differente,
che influisce sulla sua composizione e sulla sua struttura. Ad esempio, lo
splacnocranio deriva dall’ectomesenchima (ossia da cellule derivate dalla
cresta neurale), mentre il neurocranio deriva dal mesoderma (ossia cellule
derivate dai somiti). (Se questi nomi non vi dicono niente, tranquilli, saranno le prossime parole del nostro dizionario).
Nei vertebrati moderni,
l’endoscheletro, inizialmente composto da cartilagine durante la fase embrionale, può essere composto o da cartilagine (lamprede e condritti) oppure essere
ossificato (negli osteitti). Il processo di ossificazione può derivare da
invasione di tessuto osseo all’interno della cartilagine (si ottiene così un
tipo di tessuto osseo cellulare chiamato endocondrale) oppure dalla produzione di tessuto
osseo da parte dello strato più superficiale della cartilagine (osso
pericondrale, acellulare). Entrambi i processi possono avvenire simultaneamente, ma siccome
sappiamo che tessuto pericondrale è presente negli osteostraci, possiamo
identificare questo tipo di ossificazione come la prima ad essere apparsa, tra
le due.
Oltre a ciò, varie forme
fossili posseggono particolari tipi di tessuti dell’endoscheletro, di forma e
composizione diagnostica, che non si ritrovano in altri taxa e che quindi sono
tipici di singoli gruppi e possono aiutarci anche nell’indagine filogenetica.
Un esempio è la galeaspidina, che si trova solo ed esclusivamente
nell’endoscheletro dei galeaspidi (Wang et al., 2005).
A questo punto però bisogna
fare una precisazione: il fatto che lamprede e condritti possiedono entrambi
uno scheletro cartilagineo potrebbe far pensare che questa sia la condizione
primitiva per gnatostomata (e che quindi gli squali siano in qualche modo primitivi
rispetto agli ostetti). Tuttavia, se guardiamo nel record fossile, vediamo che
molti stem gnatostomi possiedono un endoscheletro ossificato, formato inizialmente da tessuto osso pericondrale. Il tessuto endocondrale compare negli osteitti e negli acantodi.
Dunque, si può ipotizzare
che lungo la linea degli gnatostomi, l’ossificazione sia comparsa all’incirca
in forme senza mascelle come gli osteostraci, sotto forma di osso pericondrale,
che dopo la comparsa delle mascelle (o around) sia comparso il tessuto
epicondrale e che lo stato cartilagineo dei condritti sia una caratteristica
derivata. I condritti dunque non sono cartilaginei come lo sono i ciclostomi. I
primi hanno perso tessuto osseo, i secondi non l’hanno mai sviluppato.

Per riassumere, possiamo
dire che la condizione primitiva del più recente antenato di crown
gnathostomata consisteva in un esoscheletro con condizione macromerica, formato
da dentina, enameloid e tessuto membrano, e in un endoscheletro con
ossificazione sia endocondrale che pericondrale.
Chiudo sottolineando come,
anche questa volta, abbiamo visto come alcune caratteristiche che sono generalmente
ritenute primitive se consideriamo solo quello che vediamo oggi, come lo
scheletro cartilagineo degli squali, se messe all’interno di un contesto che
prenda in considerazione l’intera storia evolutiva dei vertebrati e le prove
fossili possono avere un significato ben diverso, passando a volte da essere
primitive a derivate.
Per capire i cosa, i come e
i quando dell’evoluzione, è fondamentale una visione precisa e ampia, che tenga
in considerazione i punti di collegamento, le similitudini, le apparenze, le
convergenze e le omologie, derivate da fenomeni di mutamento che operano da
milioni di anni. Soffermarci solo su quello che è oggi per capire quello che è
stato il passato, è come voler ricostruire una ricetta basandoci sul gusto che
sentiamo da un unico assaggino.
--------------------------------------------------
Bibliografia:
--------------------------------------------------
Bibliografia:
- Janvier, P. - 1996
Early Vertebrates, Oxford University
Press, Oxford
- Wang N.,
Donoghue, P. C. J., Smith, M. M. and Sansom, I. J. - 2005
Histology of the
galeaspid dermoskeleton and endoskeleton, and the origin and early evolution of
the vertebrate cranial endoskeleton. Journal
of Vertebrate Paleontology 25 (4): 745 – 756
5 commenti:
Grande! Grazie!
Grande! Grazie!
Grazie a te per leggere il blog. purtroppo ora sono molto impegnato a finire il mio dottorato ma un giorno conto di riprendere :-) grazie comunque per la fiducia
Marco, mi piacerebbe mettermi in contatto con te. Mi lasci un riferimento dove posso scriverti privatamente?
Certo, scrivimi pure a makko.kildonan@gmail.com
Posta un commento