Le forme di vita del Cambriano sono sicuramente uno degli argomenti di maggior fascino dal punto di vista paleontologico. Sia perché esse ci appaiono così strane rispetto a quelle presenti oggi, sia perché appartengono ad un tempo estremamente lontano, quando la terraferma era disabitata e lo status chimico-fisico-ecologico del nostro pianeta era ben diverso da quello attuale.
Ma uno dei fattori che aumenta il grado di interesse e di fascino intorno a questo periodo della storia della Terra è sicuramente legato al fatto che di esso sappiamo ancora poco, spesso troppo poco.
Questo discorso ben si applica alle forme di vita cambriane che fanno parte del gruppo dei cordati, clade a cui anche noi apparteniamo insieme anche a tutti gli altri vertebrati, estinti e non (di cui ho parlato qui). Conoscere dunque i primi cordati e la loro evoluzione rappresenta un punto fondamentale per lo studio non solo dell’evoluzione della vita ma anche della storia della nostra specie.
Purtroppo, come spesso accade quando si parla di record fossile, la nostra conoscenza delle prime fasi della storia dei cordati e dei vertebrati è ancora nebbiosa e incerta, a causa dei pochi siti cambriani in cui sono stati trovati fossili di cordati, animali dal corpo molle che, appunto, non si fossilizzano con facilità.
Per nostra fortuna però, i pochi posti da cui sono noti resti di cordati cambriani presentano condizioni di conservazione straordinarie. Sto parlando dei due incredibili Lagerstatten di Burgess Shale e Chengjiang, di cui ho parlato già approfonditamente diverse volte. In essi non solo si sono conservati alcuni taxa di cordati cambriani, ma essi presentato uno stato di fossilizzazione tale che è spesso possibile analizzare con estremo dettaglio la loro morfologia.
Ed è proprio analizzando alcuni fossili di cordati provenienti da questi siti che un nuovo recente articolo, pubblicato su Nature da Conway Morris e Caron (2014), ha evidenziato l’importanza dello studio di queste forme nella nostra comprensione dell’evoluzione dei vertebrati, persino di alcune caratteristiche, come l’origine delle mascelle, che sembrano poco evidenti in animali agnati e senza scheletro.
Ma uno dei fattori che aumenta il grado di interesse e di fascino intorno a questo periodo della storia della Terra è sicuramente legato al fatto che di esso sappiamo ancora poco, spesso troppo poco.
Questo discorso ben si applica alle forme di vita cambriane che fanno parte del gruppo dei cordati, clade a cui anche noi apparteniamo insieme anche a tutti gli altri vertebrati, estinti e non (di cui ho parlato qui). Conoscere dunque i primi cordati e la loro evoluzione rappresenta un punto fondamentale per lo studio non solo dell’evoluzione della vita ma anche della storia della nostra specie.
Purtroppo, come spesso accade quando si parla di record fossile, la nostra conoscenza delle prime fasi della storia dei cordati e dei vertebrati è ancora nebbiosa e incerta, a causa dei pochi siti cambriani in cui sono stati trovati fossili di cordati, animali dal corpo molle che, appunto, non si fossilizzano con facilità.
Per nostra fortuna però, i pochi posti da cui sono noti resti di cordati cambriani presentano condizioni di conservazione straordinarie. Sto parlando dei due incredibili Lagerstatten di Burgess Shale e Chengjiang, di cui ho parlato già approfonditamente diverse volte. In essi non solo si sono conservati alcuni taxa di cordati cambriani, ma essi presentato uno stato di fossilizzazione tale che è spesso possibile analizzare con estremo dettaglio la loro morfologia.
Ed è proprio analizzando alcuni fossili di cordati provenienti da questi siti che un nuovo recente articolo, pubblicato su Nature da Conway Morris e Caron (2014), ha evidenziato l’importanza dello studio di queste forme nella nostra comprensione dell’evoluzione dei vertebrati, persino di alcune caratteristiche, come l’origine delle mascelle, che sembrano poco evidenti in animali agnati e senza scheletro.
Conwey Morris e Caron ridescrivono Metaspriggina walcotti (Simonetta and Insom, 1993) sulla base di almeno 100 nuovi esemplari provenienti da Burgess Shale e dal sito coevo di Marble Canyon, entrambi in Canada e risalenti al Cambriano medio, circa 505 milioni di anni fa. Gli esemplari sono conservati su lastra, quindi in due dimensioni, ma il grado di conservazione è molto buono, con evidenza anche di alcuni organi interni. La grandezza media è di circa 6 centimetri di lunghezza e 1 centimetro di altezza nel punto più alto.
Grazie al riesame di questi nuovi fossili, oggi possiamo avere un’idea molto più chiara di cosa sia Metaspriggina.
Grazie al riesame di questi nuovi fossili, oggi possiamo avere un’idea molto più chiara di cosa sia Metaspriggina.
 |
| Esemplare fossile di Metaspriggina. Potete notare l'alto grado di conservazione che permette uno studio in dettaglio del fossile (da Conway Morris and Caron, 2014). |
Esso si presenta come un animale dal corpo fusiforme e compresso lateralmente, più alto nella parte centrale e che man mano si restringe verso la parte terminale del corpo, con una testa non separata dal resto del corpo in cui si distinguono non senza difficoltà i piccoli occhi e l’apertura buccale arrotondata. Manca qualsiasi segno di pinne anche se, considerando gli altri cordati cambriani come Haikouichthys, possiamo ipotizzare che anche Metaspriggina presentasse almeno una pinna dorsale e una ventrale lungo la parte finale del corpo, probabilmente non conservata per motivi tafonomici. Una struttura cilindrica, rappresentante la notocorda, è visibile a partire dalla testa fino alla punta della coda.
Dal punto di vista filogenetico, Metaspirggina è considerato un cordato vicino al gruppo dei vertebrati. Nel cladogramma presentato da Conwey-Morris and Caron esso forma una politomia insieme a Millokinmingia, Haikouichthys e Vertebrata.
 |
| Cladogramma che mostra la posizione di Metaspriggina all'interno della filogenesi dei cordati (da Conway Morris and Caron, 2014) |
Sebbene ad uno sguardo superficialmente esso mostra tutto ciò che ci aspetteremmo di trovare in un cordato cambriano, esaminandolo più a fondo, però, questo taxon presenta dei dettagli anatomici molto particolari che ci consentono di ipotizzare alcuni aspetti delle sua ecologia prima sconosciuti.
Ad esempio, grazie al perfetto stato di fossilizzazione, è possibile osservarne la struttura dei miomeri (fasci muscolari) e comparandola con quella di altri cordati acquatici è possibile ipotizzare ad esempio che Metaspriggina fosse un nuotatore attivo abbastanza efficace e rapido nei movimenti. A ciò è presumibilmente legata la morfologia dei suoi occhi, che per la prima volta possono essere ricostruiti come aventi una struttura abbastanza complessa con il possesso di una lente. In mezzo ad essi si trovano un paio di sacche nasali, probabilmente collegate ad un singolo dotto nasale. Inoltre, la testa è in qualche modo separata dal resto del corpo e sembra possedere qualche capacità di torsione. Dunque, in generale possiamo dire che Metaspriggina era un animale più attivo di quanto si pensasse prima, capace di nuotare con efficienza e di interagire con l’ambiente in maniera complessa.
(Nota: nonostante gli autori descrivano questo dettaglio anatomico senza fare alcun commento, la presenza di narici pari sarebbe un carattere importantissimo. Infatti, narici esterne pari sono presenti infatti considerate una caratteristica distintiva dei vertebrati con mascelle, e dunque trovarne evidenza in un cordato non vertebrato e agnato sarebbe un dato assolutamente importante. Purtroppo però gli autori sembrano soprassedere su questo. Indagherò)
Ad esempio, grazie al perfetto stato di fossilizzazione, è possibile osservarne la struttura dei miomeri (fasci muscolari) e comparandola con quella di altri cordati acquatici è possibile ipotizzare ad esempio che Metaspriggina fosse un nuotatore attivo abbastanza efficace e rapido nei movimenti. A ciò è presumibilmente legata la morfologia dei suoi occhi, che per la prima volta possono essere ricostruiti come aventi una struttura abbastanza complessa con il possesso di una lente. In mezzo ad essi si trovano un paio di sacche nasali, probabilmente collegate ad un singolo dotto nasale. Inoltre, la testa è in qualche modo separata dal resto del corpo e sembra possedere qualche capacità di torsione. Dunque, in generale possiamo dire che Metaspriggina era un animale più attivo di quanto si pensasse prima, capace di nuotare con efficienza e di interagire con l’ambiente in maniera complessa.
(Nota: nonostante gli autori descrivano questo dettaglio anatomico senza fare alcun commento, la presenza di narici pari sarebbe un carattere importantissimo. Infatti, narici esterne pari sono presenti infatti considerate una caratteristica distintiva dei vertebrati con mascelle, e dunque trovarne evidenza in un cordato non vertebrato e agnato sarebbe un dato assolutamente importante. Purtroppo però gli autori sembrano soprassedere su questo. Indagherò)
 |
| Particolare degli occhi e delle narici pari di un esemplare di Metaspriggina (da Conway Morris and Caron, 2014) |
L’alto grado di conservazione del fossile consente anche di ricostruire alcuni aspetti della sua morfologia interna, come lo stomaco, l’esofago e quello che potrebbe essere il cuore, ma le evidenze a riguardo non permettono un’analisi in dettaglio di queste strutture.
La zona anatomica più significativa di questo cordato è però quella centrale, in cui risiedono le branchie.
In quest’area Metaspriggina mostra due serie di barre cartilaginee, una sopra all’altra, sette per parte per un totale di 14 barre per lato e 28 in totale. La fila superiore di barre cartilaginee corre parallelamente alla fila inferiore e, in visione laterale, esse sono ricurve ventralmente, proprio come negli archi branchiali degli altri cordati. Associate alle barre, tranne che al primo paio (barra dorsale+ventrale sinistra e barra dorsale+ventrale destra) si trovano i filamenti che costituivano le superfici di scambio dell’organo branchiale.
La zona anatomica più significativa di questo cordato è però quella centrale, in cui risiedono le branchie.
In quest’area Metaspriggina mostra due serie di barre cartilaginee, una sopra all’altra, sette per parte per un totale di 14 barre per lato e 28 in totale. La fila superiore di barre cartilaginee corre parallelamente alla fila inferiore e, in visione laterale, esse sono ricurve ventralmente, proprio come negli archi branchiali degli altri cordati. Associate alle barre, tranne che al primo paio (barra dorsale+ventrale sinistra e barra dorsale+ventrale destra) si trovano i filamenti che costituivano le superfici di scambio dell’organo branchiale.
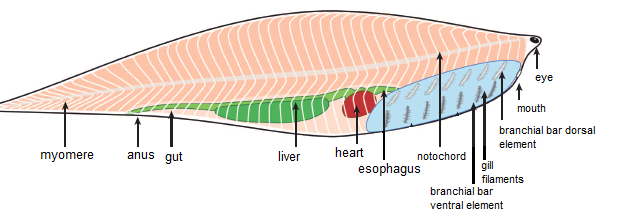 |
| Anatomia interna di Metaspirggina (modificato da Conway Morris and Caron, 2014) |
La presenza di queste barre bipartite nella regione branchiale di Metaspriggina e la loro struttura anatomica sono particolarmente importanti poiché richiamano la morfologia degli archi branchiali degli gnatostomi.
Ad esempio, negli gnathostomi non tutti gli archi branchiali sono associati alle branchie, ma esse si originano a partire dal secondo (arco hyoideo), mentre il primo arco (arco mandibolare) è legato alla bocca e, nei vertebrati con mascelle, agli elementi che poi costituiscono la maggior parte di mascella e mandibola. Metaspriggina presenta una serie di archi branchiali il cui primo però, più robusto, non sembra associato ai filamenti branchiali che invece sono presente vicino agli altri archi. Sebbene, come dicono gli autori dell’articolo, l’omologia tra il primo arco branchiale di Metaspriggina e il primo arco branchiale degli gnathostomi sia ancora tutta da verificare, questa somiglianza è interessante e, magari associata ad ulteriori futuri ritrovamenti, potrebbe essere altamente informativa per lo studio dell’origine delle mascelle.
Inoltre, la presenza di due paia di serie di archi branchiali paralleli in Metaspriggina ricorda la struttura presente negli gnathostomi (come abbiamo visto qui), dove gli archi branchiali sono formati, ciascuno, da diversi elementi bastoncellari (ceratoiale, epiiale, ioidale, etc..). La condizione bipartita degli archi branchiali è considerata da sempre una caratteristica comune degli gnathostomi, rappresentante dunque la condizione originaria per questo gruppo. I cyclostomi invece, possiedono archi branchiali non bipartiti ma con i vari elementi fusi insieme.
Secondo Conway Morris e Caron, le due barre (per ogni segmento) presenti in Metaspriggina sarebbero omologhe agli elementi degli archi branchiali degli gnatostomi, rispettivamente all’epibranchiale e al ceratobranchiale. Considerare omologi questi elementi in Metaspriggina e negli gnathostomi implicherebbe sia che archi branchiali bipartiti non solo sono primitivi negli gnathostomi ma anche potrebbero essere la condizione standard dei vertebrati, sia la struttura delle branchie dei cyclostomi, così diversa da quella di tutti gli altri vertebrati, è in realtà una peculiarità evolutiva di questo gruppo e non retaggio ancestrale.
I cyclostomi non possiedonoinfatti archi branchiali bipartite ma le branchie sono sostenute da un complessa rete di barre cartilaginee, verticali e orizzontali, a formare quello che viene definito “sacco branchiale”.
Ad esempio, negli gnathostomi non tutti gli archi branchiali sono associati alle branchie, ma esse si originano a partire dal secondo (arco hyoideo), mentre il primo arco (arco mandibolare) è legato alla bocca e, nei vertebrati con mascelle, agli elementi che poi costituiscono la maggior parte di mascella e mandibola. Metaspriggina presenta una serie di archi branchiali il cui primo però, più robusto, non sembra associato ai filamenti branchiali che invece sono presente vicino agli altri archi. Sebbene, come dicono gli autori dell’articolo, l’omologia tra il primo arco branchiale di Metaspriggina e il primo arco branchiale degli gnathostomi sia ancora tutta da verificare, questa somiglianza è interessante e, magari associata ad ulteriori futuri ritrovamenti, potrebbe essere altamente informativa per lo studio dell’origine delle mascelle.
Inoltre, la presenza di due paia di serie di archi branchiali paralleli in Metaspriggina ricorda la struttura presente negli gnathostomi (come abbiamo visto qui), dove gli archi branchiali sono formati, ciascuno, da diversi elementi bastoncellari (ceratoiale, epiiale, ioidale, etc..). La condizione bipartita degli archi branchiali è considerata da sempre una caratteristica comune degli gnathostomi, rappresentante dunque la condizione originaria per questo gruppo. I cyclostomi invece, possiedono archi branchiali non bipartiti ma con i vari elementi fusi insieme.
Secondo Conway Morris e Caron, le due barre (per ogni segmento) presenti in Metaspriggina sarebbero omologhe agli elementi degli archi branchiali degli gnatostomi, rispettivamente all’epibranchiale e al ceratobranchiale. Considerare omologi questi elementi in Metaspriggina e negli gnathostomi implicherebbe sia che archi branchiali bipartiti non solo sono primitivi negli gnathostomi ma anche potrebbero essere la condizione standard dei vertebrati, sia la struttura delle branchie dei cyclostomi, così diversa da quella di tutti gli altri vertebrati, è in realtà una peculiarità evolutiva di questo gruppo e non retaggio ancestrale.
I cyclostomi non possiedonoinfatti archi branchiali bipartite ma le branchie sono sostenute da un complessa rete di barre cartilaginee, verticali e orizzontali, a formare quello che viene definito “sacco branchiale”.
Le fessure branchiali non giacciono tra una barra e l’altra, come negli gnathostomi, ma invece esternamente al sacco branchiale.
Uno stadio “intermedio” tra la condizione bipartita degli gnathostomi e la rete dei cyclostomi è visibile nell’enigmatico stem gnathostomo Euphanerops (di cui ho parlato in dettaglio qui e qui), taxon la cui posizione filogenetica è ancora tutt’altro che risolta e che presenta un intrigante mix di caratteri tra lamprede e alcuni stem gnathostomi agnati (vedere post linkati).
Oltre all’elevato numero di aperture branchiali (come i cyclostomi e diversamente dagli gnathostomi) gli archi branchiali sono fusi in una singola unità, di forma sinuosa, associata con una serie di elementi rigidi di forma stretta e ricurva (tipo “costoline”). Questo tipo di apparato branchiale, unico di questo animale, ricorda i qualche modo la “sacca branchiale” delle lamprede. Interessante suggestione data dall’analisi filogenetica presentata nell’articolo di Metaspriggina vede Euphanerops come sister-taxon dei cyclostomi. Questo potrebbe suggerire come l’anatomia branchiale di Euphanerops rappresenti la fase precedente alla profonda riorganizzazione del sostegno delle branchie presente nei cyclostomi a partire da una struttura con barre verticali bipartite, ulteriore conferma di come la morfologia dei cyclostomi, che a noi oggi appare peculiare e che viene sempre descritta come “primitivamente strana”, sia in realtà il frutto di un lungo processo evolutivo occorso dopo la separazione tra cyclostomi e gnathostomi.
Metaspriggina non solo ci presenta dunque nuovi squisiti dettaglia dell’anatomia delle bizzarre forme di vita cambriane, ma, ancora una volta, ci mostra come senza i fossili la nostra visione dell’evoluzione, se fosse basata solo sulle forme di vita che abitano intorno a noi nel nostro piccolo istante temporale, sarebbe molto, molto distorta.
P.S. W i cyclostomi ;-)
Uno stadio “intermedio” tra la condizione bipartita degli gnathostomi e la rete dei cyclostomi è visibile nell’enigmatico stem gnathostomo Euphanerops (di cui ho parlato in dettaglio qui e qui), taxon la cui posizione filogenetica è ancora tutt’altro che risolta e che presenta un intrigante mix di caratteri tra lamprede e alcuni stem gnathostomi agnati (vedere post linkati).
Oltre all’elevato numero di aperture branchiali (come i cyclostomi e diversamente dagli gnathostomi) gli archi branchiali sono fusi in una singola unità, di forma sinuosa, associata con una serie di elementi rigidi di forma stretta e ricurva (tipo “costoline”). Questo tipo di apparato branchiale, unico di questo animale, ricorda i qualche modo la “sacca branchiale” delle lamprede. Interessante suggestione data dall’analisi filogenetica presentata nell’articolo di Metaspriggina vede Euphanerops come sister-taxon dei cyclostomi. Questo potrebbe suggerire come l’anatomia branchiale di Euphanerops rappresenti la fase precedente alla profonda riorganizzazione del sostegno delle branchie presente nei cyclostomi a partire da una struttura con barre verticali bipartite, ulteriore conferma di come la morfologia dei cyclostomi, che a noi oggi appare peculiare e che viene sempre descritta come “primitivamente strana”, sia in realtà il frutto di un lungo processo evolutivo occorso dopo la separazione tra cyclostomi e gnathostomi.
Metaspriggina non solo ci presenta dunque nuovi squisiti dettaglia dell’anatomia delle bizzarre forme di vita cambriane, ma, ancora una volta, ci mostra come senza i fossili la nostra visione dell’evoluzione, se fosse basata solo sulle forme di vita che abitano intorno a noi nel nostro piccolo istante temporale, sarebbe molto, molto distorta.
P.S. W i cyclostomi ;-)
-----------------------------------------------------------------
Bibliografia:
- Conway Morris S. and Caron J-B. (2014)
A primitive fish fromthe Cambrian of North America
Nature, doi:10.1038/nature13414
1 commento:
Ho imparato tanto da questo blog che é riuscito un pochino a scrostare il muro della mia ignoranza settoriale, ma, negli ultimi tempi mi sono ripetutamente stupito per gli studi che hanno cambiato le prospettive: prima Entelognatus suggerisce che lo scheletro macromerico degli osteitti è basale e quello dei condritti é derivato. Poi Ozarcus sembra ribadire la medesima cosa riguardo la disposizione delle ossa degli archi branchiali. Infine Metaspriggina pare indicarci che i cyclostomi abbiano in realtà un mucchio di caratteristiche derivate rispetto ad una disposizione basale degli archi branchiali molto più simile a quella dei gnatostomi stem. Bello.
Posta un commento