Questa estate avevo pubblicato un mini paleoquiz.
Scusa se ci ho messo più tempo del previsto, dovevo prima verificare alcune cose.
La domanda era: cos'hanno in comune il teropode cretaceo Struthiomimus e un odierno camaleonte?


Il titolo svela in parte la risposta. La struttura che
accomuna i due animali è la mano e, aggiungo ora, la
conformazione del cinto pettorale.
I camaleonti sono animali molto particolari, sia dal
punto di vista etologico, visto il comportamento complesso legato al
corteggiamento e a tutta una serie di interazioni tra individui basate sulla
variazione di colore e tonalità della pelle, sia dal punto di vista puramente
anatomico.
Lo scheletro del camaleonte presenta numerosi tratti
anatomici legati ad un perfetto adattamento per uno stile di vita arboricolo:
gli arti sono posizioni quasi in maniera parasaggitale (in maniera simile a
dinosauri, uccelli e mammiferi) o comunque molto meno "sprawling" (se
qualcuno ha un termine corretto per tradurre questa parola in italiano, lo
ringrazio davvero) rispetto alla normale postura dei rettili, anche di quelli
arboricoli.
Sauri arboricoli come le lucertole del genere Anolis, Dypsosaurus,
Agama, e varie iguane e gechi, posseggono comunque arti che si protraggono dal
corpo lateralmente (appunto, secondo postura detta "sprawling").
La postura eretta del camaleonte è particolarmente
pronunciata negli arti anteriori, mentre quelli posteriori sono più simili agli
altri rettili arboricoli (Fisher et al., 2010)
Ma del cinto pettorale, degli arti anteriori e della
"mano" dei camaleonti, l'argomento centrale di questo post, voglio
parlare dopo.

Gli altri adattamenti legati ad uno stile di vita
arboricolo presenti nei camaleonti sono una coda prensile (anche se non tutti i
camaleonti utilizzano la coda per svolgere questo ruolo, e alcuni non sono in
grado di sostenersi facendo affidamento in prevalenza sulla coda), un corpo
compresso lateralmente (caratteristica dei camaleonti che li differenzia dagli
arti rettili arboricoli, di solito compressi dorsoventralmente o comunque con
conformazione corporea classica da sauro), grossi occhi, che possono essere
mossi separatamente e che forniscono ai camaleonti una visione stereoscopica,
per vedere bene anche nelle zone di poca luce come possono essere le fronde
degli alberi e cacciare animali rapidi come gli insetti volatori, e
soprattutto, parlando di camaleonti, la modificazione dell'osso ioideo tale da
poter sorreggere una lunga e potente lingua muscolosa, che termina con una
porzione appiccicosa. I camaleonti utilizzano la loro lingua per tendere
agguati alle loro prede.

Se guardiamo con maggior dettaglio lo scheletro del
camaleonte, quello che si può notare subito e che veramente distingue questo
animale dagli altri tetrapodi è la conformazione del suo cinto pettorale.
Nella sua forma più completa, il cinto pettorale è
formato da un cinto primario e un cinto secondario (Peterson, 1973; Nicholls
and Russell, 1985). Il cinto primario è composto dalla scapola e dal coracoide,
ossa pari che solitamente sono strettamente connessi a formare un complesso
funzionale molto importante. Scapola e coracoide non sono connessi alla colonna
vertebrale (a differenza del cinto pelvico, il cinto pettorale non è connesso
direttamente alla colonna, se non per mezzo di legamenti e muscoli) e a queste
due ossa sono generalmente collegate le modifiche morfologiche legati ai vari
adattamenti locomotori riguardanti il cinto pettorale.
Il cinto secondario è formato dalle clavicole (pari),
da un interclavicola impari (nei rettili non uccelli e anfibi) e da uno sterno
che può essere pari (nei rettili non uccelli) o impari (negli uccelli).
Il cinto secondario rende più stabile il cinto
pettorale, evitando di dislocare in maniera eccessiva le articolazioni (a volte
limitandone però i movimenti). Vari adattamenti
Il cinto secondario serve a rendere più stabile il
cinto primario e in generale tutto il complesso del cinto pettorale, evitando
un eccessivo dislocamento delle articolazioni, anche se spesso ciò limita
parzialmente i movimenti.
 |
| Cinto pettorale di coccodrillo, come esempio del tipico cinto pettorale dei rettili. i c. = inerclavicola co.= coracoide sc.= scapola. |
A volte, per precisi adattamenti anatomici, il cinto
secondario può essere fortemente modificato, ridotto, eliminato.
E questo è il caso dei camaleonti.
Dato il loro estremo adattamento ad uno stile di vita
arboricolo, i camaleonti hanno un cinto pettorale fortemente modificato
rispetto allo standard degli altri tetrapodi.
Dovendo spostarsi sugli alberi, essi devono fare in
modo da favorire il più possibile la brachiazione e di rendere il cinto
pettorale il più mobile possibile.
I camaleonti hanno risolto questo problema riducendo
fortemente il cinto secondario (non ci sono ne clavicole ne interclavicole),
tale che il loro cinto è formato solo da scapola, coracoide e sterno.
Ma c'è' di più: scapola e coracoide sono fusi insieme
in un robusto complesso scapolacoracoideo, mentre lo sterno è relativamente
ridotto, non ampio e piato come nella maggior parte degli altri animali, ma
compresso lateralmente a formare una sorta di V.
Queste modificazioni ossee permettono un particolare
tipo di movimento delle ossa del cinto, che rappresenta la vera novità dei
camaleonti. Infatti, durante la brachiazione, i due complessi scapolacoracoidei
possono muoversi in maniera indipendente, scorrendo all'interno di una fossa
situata sulla faccetta laterale esterna dello sterno. In questo modo la
possibilità di estendere le braccia in due direzioni opposte (ad esempio, una
protesa in avanti e una protesa all'indietro) è portata all'estremo, in modo che, grazie a tale escursione delle braccia, i
camaleonti possono facilmente spostarsi su superfici discontinue anche
abbastanza intricate.
Ciò è
possibile anche grazie all'assenza dell'inteclavicola, la cui presenza blocca i due
coracoidi.
 |
| Cinto pettorale di camaleonte in visione laterale, anteriore e ventrale. |
Ma, vi starete giustamente chiedendo, cosa c'entra un
animale così specializzato a vivere sugli alberi con Struthiomimus, un
dinosauro, bipede, corridore, che ovviamente non viveva sugli alberi e che
sembra non avere niente a che fare, almeno dal punto di vista ecologico, con il
camaleonte?
Un attimo e ci arriviamo.
Struthiomimus (Osborn, 1917) è un teropode del cretaceo superiore nordamericano appartenente al gruppo degli ornithomimosauri. Essi si distinguono per il cranio "simile" a quello degli uccelli (cranio piccolo, rostro allungato, presenza di becco senza denti, orbite grandi), corpo slanciato, collo relativamente lungo, arti anteriori snelli e lunghi, con mani munite di dita prensili, arti posteriori con struttura metatarsale e adattati alla corsa (per maggiori informazioni, consultare una fonte migliore di questa. Paleostories non è un blog specialistico sui dinosauri).
Pur essendo complessivamente molto diverso per
le dimensioni, la postura bipede e l’aspetto generale, Struthiomimus presenta alcuni aspetti che possono collegarlo al
nostro bizzarro arboricolo.
 |
| Disegno dello scheletro di Struthiomimus. |
 |
| Cinto pettorale di Struthiomimus. |
Nicholls e Russell, in uno studio di dettaglio sul
cinto di Struthiomimus (1985), fanno addirittura notare come “in effetti, il
cinto pettorale di Chamaeleo ricorda così tanto la condizione tipica dei
teropodi che l’unica ovvia differenza è data dalle dimensioni”
Ma una somiglianza ancora più stretta è presente tra i
camaleonti e Struthiomimus: come in questi rettili attuali, in
Struthiomimus il cinto è modificato tale da presentare un cinto secondario
molto ridotto. Come nel camaleonte, la scapola di Struthiomimus è
piuttosto allungata e poco espansa dorsalmente. Le clavicole sono assenti, così
come l'interclavicola (anche se queste mancanze potrebbero essere il risultato
di cause tafonomiche), e lo sterno è meno piatto e sviluppato che in altri
dinosauri, e articolato in maniera tale da risultare quasi in linea con il
coracoide (questa è una differenza sostanziale con il camaleonte, ma il
risultato funzionale è comunque simile).
Nicholls e Russel (1985) interpretano queste
modificazioni come indice di una mobilità elevata per quanto riguarda il cinto
pettorale di questo animale.
Ma le similitudini non si fermano qui.
La maggior parte dei rettili arboricoli attuali, come
Anolis, le iguane o i gechi, hanno una struttura della “mano” non particolarmente
modificata, con in genere mani e piedi simili, piatti, spesso con dita larghe e
corte (gechi) oppure con dita di varia lunghezza, sottili e distanziate l'una dall'altra.
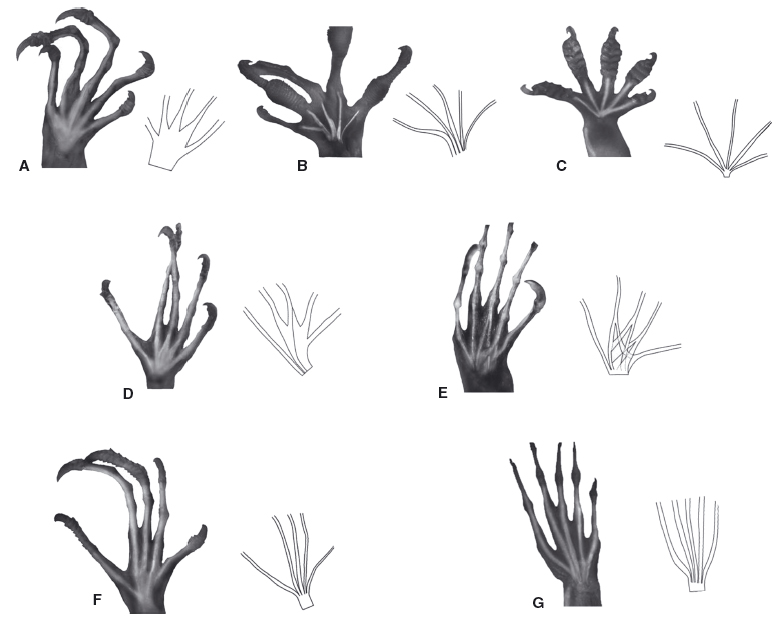 |
| Esempi di mani di varie specie di Anolis |
Gli
arti dei camaleonti sono invece molto specializzati: mani i piedi possiedono dita,
della stessa lunghezza, unite a formare due gruppi funzionali indipendenti, in
una forma simile ad una pinza, con due dita in una parte e tre dita nell’altra.
Nella mano l’unità più esterna è formata da due dita, la più interna da due. Nel
piede troviamo il contrario.
Le dita sono racchiuse in una membrana di pelle che si
estende fino alla base della falange ungueale, facendo spuntare le unghie solo
per una piccola porzione. La membrana ha anche la funzione di tenere le dita
unite, parallele e molto vicine, in modo tale da farle funzionare come una
singola unità.
Le dita, di uguale lunghezza, sono racchiuse da una
membrana di pelle che si estende fino alla base degli ungueali, tenendo le dita
parallele e molto vicine, facendole operare appunto come una singola unità.
 |
| Mano di camaleonte durante l'atto di presa |
Polsi e caviglie sono molto mobili e dotati di
un’ampia escursione rotatoria.
Queste modificazioni di mani e piedi donano ai
camaleonti un’ottima capacità di presa, atta a muoversi su superfici irregolari
e sottili. La conformazione della mano è fortemente modificata per far si ce
essa possa essere un organo ottimamente funzionale nel muoversi su superfici
irregolari e sottili. Questa specializzazione dei loro arti è così radicata nel
gruppo dei camaleonti che anche le (poche) specie terrestri hanno la stessa
struttura a pinza, pur non vivendo sugli alberi.
Anche se il polso non è così mobile come quello dei
camaleonti, la mano di Struthiomimus presenta alcune importanti similitudini
con quella di questi rettili arboricoli (Nicholls and Russell, 1985).
 |
| Mano di Struthiomimus |
Per prima cosa, la mano di Struthiomimus non presenta
le tipiche caratteristiche della mano raptatoria di molti altri dinosauri, come
ad esempio vari allosauroidi, deinonychosauri e coelurosauridi. In una tipica
mano raptatoria, le dita II e III sono diseguali in lunghezza e abbastanza
distanti tra di loro, in modo tale che quando le dita vengono estese si aumenta
la superficie di presa della mano.
Inoltre, spesso il polso è capace di ampi movimenti rotatori e il dito I
è opponibile o semi-opponibile.
Una sostanziale caratteristica che distingue questo
dinosauro dagli altri dinosauri (e anche dagli altri ornithomimosauri) è il
possesso di dita della stessa lunghezza. Queste dita (la sua mano è fornita di
tre dita, tutte e tre funzionali e con ungueale) sono posizionate in maniera
tale che il II e il III dito sono posti in stretto legame tra di loro, mentre
il I è sostanzialmente più distante.
 Il primo I non è opponibile (Ostrom 1969, Nicholls and
Russel, 1985), il polso è piuttosto rigido e nel momento di massima estensione,
le dita II e III sono posizionate in parallelo, una vicina all’altra, mentre il
dito I diverge da essi. Questo perché la
superficie articolare distale del metacarpale I è ruotata in senso antiorario
rispetto la linea mediana della mano. Nei theropodi con mano raptatoria,
invece, la superficie articolare distale del I dito è ruotata in senso orario,
in modo tale che quando la mano viene iperestesa tutte e tre le dita
convergono (figura a sinistra).
Il primo I non è opponibile (Ostrom 1969, Nicholls and
Russel, 1985), il polso è piuttosto rigido e nel momento di massima estensione,
le dita II e III sono posizionate in parallelo, una vicina all’altra, mentre il
dito I diverge da essi. Questo perché la
superficie articolare distale del metacarpale I è ruotata in senso antiorario
rispetto la linea mediana della mano. Nei theropodi con mano raptatoria,
invece, la superficie articolare distale del I dito è ruotata in senso orario,
in modo tale che quando la mano viene iperestesa tutte e tre le dita
convergono (figura a sinistra).
In Struthiomimus, le falangi ungueali potevano essere
molto flesse, e questo, insieme alla struttura rigida del polso e alle dita
posizionate come se (come avviene nei camaleonti) fossero due diverse unità
funzionali (dito I semi opponibile vs dito II + dito III) permettono di fare
paragoni con la struttura delle mani di altri animali attuali.
Nicholls and Russell (1985) ipotizzano addirittura che
le due dita funzionalmente interconnesse (dito II e III) fossero racchiuse
insieme in una membrana di pelle, proprio come avviene oggi nei camaleonti e nei
bradipi.
La mano di Struthiomimus possiede infatti caratteristiche
tali da farne un’ottima pinza o gancio, in maniera simile a quanto avviene nei
bradipi, nei formichieri e, appunto, nei camaleonti (anche se nella mano dei
camaleonti la modifica per una mano “a pinza” è ancora più estrema).
Come possiamo dunque collegare questa somiglianza tra
le mani di animali così diversi?
Struthiomimus e gli altri ornithomimosauri sono dotati
di un becco piatto e senza denti (con eccezioni) e dunque è ipotizzabile che
fossero erbivori o al massimo onnivori.
Il possesso di un cinto molto mobile, dato dalla forte
riduzione del cinto secondario che rendeva meno stabile il cinto primario, è
correlata ad un’elevata mobilità delle braccia.
Questo si aggiunge ad una mano con una conformazione
molto particolare, a gancio o a pinza, con due dita da una parte e un dito,
divergente, dall’altra.
Secondo Nicholls e Russell (1985) questa morfologia
può essere collegata ad un preciso tipo di adattamento legato ad uno stile
alimentare vegetariano, in cui la mano di Struthiomimus
potesse essere utilizzata non per catturare le prede, come nei theropodi con
mano raptatoria, ma invece per agganciare e manipolare i rami degli alberi, in
modo da avvicinarli alla bocca.
Analisi palinologiche relative alla Judith River
Formation (Jarzen, 1982) indicano la presenza di numerose felci (Polypodiaceae)
e felci arboree (Cyatheaceae e Dicksoniaceae), così come di Cycas
(Cycadophyta).
Nicholls and Russell suggeriscono che l’ampia mobilità
degli arti di Struthiomimus e la struttura a gancio delle sue mani potevano
essere utilizzate per agganciare e tirare a se le fronde di queste piante, in
modo da portarli più vicini alla bocca. La presenza di un collo lungo forse
potrebbe avvalorare questa tesi, giacché potrebbe essere correlata ad un tipo
di foraggiamento che richiede di raggiungere cibo ad un’altezza rilevante dal
suolo.
Così come il camaleonte usa le sue mani a pinza e il
suo cinto molto (molto!) mobile per raggiungere anche i rami più lontani e
muoversi con più disinvoltura sugli intricati, e spesso molto fini e
irregolari, rami degli alberi delle foreste in cui vive, così le modifiche
strutturali del cinto e della mano di Struthiomimus
miglioravano la sua interazione con l’ambiente circostante.
 |
| Mano di Struthiomimus, con dita flesse |
 |
| Fonte di ispirazione? |
Non è la prima volta che animali molto distanti
filogeneticamente evolvono strutture simili per compiere azioni che richiedono
una simile biomeccanica.
Ovviamente, non sapremo mai (forse) se la mano di Struthiomimus fosse davvero così simile
a quella di un camaleonte o di un brado, addirittura con una membrana tra le
dita come ipotizzato da Nicholls e Russell (1985). Mi piace però scoprire come certi
dettagli di animali apparentemente molto noti sfuggono spesso alle nostre
menti.
In un mondo in cui si disegnano le più assurde e
curiose strutture anatomiche agli animali preistorici, sull’onda del “tutto è
possibile visto che conosciamo troppo poco”, perché non ho trovato alcuna
immagine di uno Struthiomimus con
mano a pinza, con membrana, mentre raccoglie una fronda di felce arborea e la
porta alla bocca? Troppo bizzarro? Poco popolare?
Se qualcuno vuole colmare questa lacuna, sarò ben
felice di esporre il suo disegno qui, su Paleostories.
----------------------------------------------------------------------------
Bibliografia:
- Fischer M. S., Krause C., Lilje K. E. 2010
Evolution of chameleon locomotion, or how to become arboreal as a reptile. Zoology 113: 67–74
- Jarzen D. M. 1982
Palynology of Dinosaur Provincial Park (Campanian) Alberta. Syllogeus 38: 1-69
- Osborn H. F. 1917
Skeletal adaptations of Ornitholestes, Struthiomimus and Tyrannosaurus. Bulletin of American Museum of Natural History 35: 733 - 771
- Nicholls
E. L. and Russell A. P. 1985
Structure and function of the pectoral girdle
and forelimb of Struthiomimus altus
(Theropoda: Ornithomimidae). Palaeontology 28 (4) : 643 - 677
- Peterson,
J. A. 1984
The locomotion of Chamaeleo
(Reptilia: Sauria) with particular reference to the forelimb. Journal of Zoology 202: 1 - 42
4 commenti:
Ciao Marco. Non ci arrivo neppure dopo il tuo aiuto, poiché continuano a sembrarmi animali con specializzazioni prensorie della mano completamente diverse. L'unica cosa che mi ha fatto pensare é un abstract in cui si dice che la specializzazione arboricola del camaleonte ha, di fatto, soppresso l'ondulazione laterale tipica della locomozione rettiliana basale in favore di un'avanzamento con arti posti più direttamente sotto al corpo; un po' quello che successe con gli antenati dei dinosauri. Ma riguardo la mano non riesco a trovare nulla. Ciao, bentornato.
Scusa Marco, ho avuto un'idea. Riguarda il fatto che i teropodi, non potendo pronare e supinare l'avambraccio, tenevano la mano col palmo rivolto medialmente? E magari il camaleonte fa lo stesso? Spero di non proferire una fesseria....
Molto tecnico e molto interessante. Ciao
Grazie :-) scusa se non ti ho risposto nei commenti precedenti ma non volevo anticipare niente ;-)
Posta un commento